Back to Home
Page of CD3WD Project or Back to list of CD3WD Publications
 |  |  | Use of Trees by Livestock (NRI, 1994, 160 p.) |  |  | Use of Trees by Livestock: Acacia | |  | (introduction...) | | | Acknowledgements | | | Foreword | | | Genus Acacia | | | Summary | | | Description and distribution | | | (introduction...) | | | Phyllodenous Acacia species | | | Bipinnate Acacia species | | | Fodder characteristics | | | Fodder yield | | | Nutritive value | | | Anti-nutritive factors | | | (introduction...) | | | Cyanogenic glycosides | | | Phenolic compounds | | | Management | | | Alternative uses | | | References and further reading | | | Appendix 1 | | | Appendix 2 |
|
Use of Trees by Livestock (NRI, 1994, 160 p.)
Use of Trees by Livestock: Acacia
J.J. Bennison and R.T.
Paterson
Acknowledgements
© Crown copyright 1993
The Natural Resources Institute (NRI) is an internationally
recognized centre of expertise on the natural resources sector in developing
countries. It forms an integral part of the British Government's overseas aid
programme. Its principal aim is to alleviate poverty and hardship in developing
countries by increasing the productivity of their renewable natural resources.
NRI's main fields of expertise are resource assessment and farming systems,
integrated pest management, food science and crop utilization.
NRI carries out research and surveys; develops pilot-scale
plant, machinery and processes; identifies, prepares, manages and executes
projects; provides advice and training; and publishes scientific and development
material.
Short extracts of material from this publication may be
reproduced in any non-advertising, nonprofit-making context provided that the
source is acknowledged as follows:
Bennison, J.J., and Paterson, R.T. (1993) Use of Trees by
Livestock 2: Acacia. Chatham, UK: Natural Resources Institute.
Permission for commercial reproduction should be sought from:
The Head, Publications and Publicity Section, Natural Resources
Institute, Central Avenue, Chatham Maritime, Kent, ME4 4TB, United Kingdom.
No charge is made for single copies of this publication sent to
the governmental and educational establishments, research institutions and
non-profit-making organizations working in countries eligible for British
Government Aid. Free copies cannot normally be addressed to individuals by name
but only under their official titles.
Natural Resources Institute
ISBN: 0 85954 334-X
Price
£2.00LG2
This booklet has been printed on paper produced using the
residue from sugar-cane
processing.
Foreword
The importance of trees and shrubs in the feeding of animals in
the tropics and sub-tropics has long been recognized by livestock owners. In
arid areas where the growth of herbaceous plants is limited by lack of moisture,
leaves and edible twigs of trees and shrubs can constitute well over 50% of the
biomass production of rangeland. At high altitudes, tree foliage may provide
over 50% of the feed available to ruminants in the dry season, branches being
harvested and carried to the animals. Even in regions of higher rainfall where
grass supplies the major proportion of the dry matter eaten by ruminants, tree
leaves and fruits can form an important constituent of the diet, particularly
for small ruminants.
In the last two decades interest in the planting of trees as a
source of feed for livestock has been encouraged by workers in research and
development, but in contrast to the hundreds of indigenous species which are
used as fodder, attention has focussed on a limited number of introduced
species. Thus there are many publications reporting the chemical composition of
Leucaena leucocephala leaves and suggesting management strategies for
utilization of the tree for fodder, but it is more difficult to find information
on alternative genera which might be equally, or more, appropriate.
The aim of this series of publications is to bring together
published information on selected genera of trees which have the potential to
increase the supply of fodder for ruminants. Each booklet summarizes published
information on the fodder characteristics and nutritive value of one genus, with
recommendations on management strategies, where available. Further, since the
leaves of woody species frequently contain secondary compounds which may have an
anti nutritional, or toxic, effect, a separate booklet summarizes the effects of
a number of these compounds. It is hoped that the booklets will provide useful
resource material for students, research and extension workers, interested in
promoting the use of trees as a source of fodder for ruminants.
Further copies of this booklet or others in the series can be
obtained by writing to the Publications Section at the Natural Resources
Institute.
Margaret Gill
Livestock Production
Programme
Genus Acacia
Family LEGUMINOSAE Subfamily MIMOSOIDEAE Tribe ACACIEAE
Principal phyllodenous species
Acacia aneura
Acacia combagei
Acacta cortacea
Acacia
cyanophylla
Acacia dealbata
Acacia decurrens
Acacia
holosericea
Acacia mearnsii
Acacia nilotica
Acacia pendula
Acacia
salicina
Acacia tortilis
Acacia trachycarpa (syn A. aff.
Iinarioides)
Acacia tumida
Principal bipinnate species
Faidherbia albida (syn. Acacia albida)
Acacia brevispica
Acacia cornigera
Acacia drepanolobium
Acacia farnesiana
Acacia
haematoxylon (syn. A. giraffae)
Acacia hebeclada subsp. hebeclada (syn. A.
stolonifera)
Acacia lasiopetala
Acacia leucophloea
Acacia mearnsii
(syn. Racosperma mearnsii)
Acacia mellifera
Acacia mellifera subsp.
detinens
Acacia nilotica
Acacia oerfota (syn. A. nubica)
Acacia
robusta
Acacia senegal
Acacia seyal
Acacia sieberiana
Acacia
tortilis
Acacia tortilis subsp. heteracantha (syn. A.
Iikatunensis)
Summary
With the African Acac�a spp. the fruit is probably more valuable
as a feed than the foliage, because with the exception of F. albida, they tend
to shed their leaves at the time of greatest feed scarcity during the dry
season. Australian species however, retain their leaves and therefore show
greater potential as fodder trees.
The productivity responses of animals reflect both the total
intake, and the availability, of nutrients in the diet. Proximate and fibre
analyses are unreliable indicators of nutritional value because of the presence
of both cyanogenic glycosides and polyphenolics. A cautious approach should be
adopted when feeding both A. leucophloea and some of the Australian species.
In some areas of Africa, Acacia spp. are the dominant browse
trees within the natural ecosystem. Often there is no choice but to utilize the
available feeds even if they do contain anti-nutritive compounds. Consumed as
part of a wider diet, the effect of anti-nutritive factors is diluted. They only
become serious in periods of extreme feed scarcity or in situations where the
opportunity for feed selection is reduced, such as in stall feeding and
cut-and-carry systems.
Acacia is a huge genus with pantropical distribution and a
notable ability to survive in harsh environments. It has potential for use in
environmental protection and for production of fodder, fuelwood, gums, and
tannins.
Description and distribution
Acacia is a pantropical and subtropical genus with species
abundant throughout Australia, Asia, Africa and the Americas. They thrive in a
diverse range of habitats and environments. Many species are well adapted to the
semi-arid and savannah regions but equally others survive in moist forest and
riverine areas, tolerating both high pH and waterlogged soils. With such
diversity, Acacia has considerable potential in a range of livestock and
agroforestry systems. In Africa and Australia, some naturally occuring species
are important in traditional pastoral and agropastoral systems, while imported
species have become commercially accepted. Trees provide fodder and shade for
livestock, improve soil fertility through nitrogen fixation and the production
of leaf litter end stabilize veils. Acacia species provide edible fruits and
seeds, gum arabic and timber for fuel, construction and fencing.
In a large genus of 800-900 species (Allen and Allen, 1981),
classification and nomenclature can be complex, particularly as there have been
a number of revisions and attempts to subdivide the genus. Pedley (1986; 1987)
advocated that it be divided into three separate genera, namely Acacia, with 200
species best represented in Africa and South America, Senegalia, with about 150
species with the same geographical distribution, and Racosperma, with about 850
species virtually confined to Australia. While the suggestion may have merit, it
has not yet been generally accepted. This text uses the traditional nomenclature
adopted by standard flora. Where a name has been changed as a result of a
generally accepted revision, the name used in the original reference is listed
as a synonym. The names of the African species are those included in the
check-list produced by Lock (1989) as part of the programme of the International
Legume Database and Information Service (ILDIS).
One widely accepted recent change is the reclassification of
Acacia albida as Faidherbia albida, although Coe and Beentje (1991) stated that
the justification for the change was primarily phytochemical. In this
publication, for purposes of discussion and comparison, the name F. albida is
accepted but it is considered as though it had remained within the genus Acacia,
since it shares many agronomic characteristics with species that have not been
reclassified.
Various workers have divided the genus into divisions, series
and subseries (Bentham, 1864) or sections (Taubert, 1894) but for ease of
description and an understanding of the fodder potential, it is simplest to
describe Acacia spp. as being either phyllodenous (principally Australian in
origin) or bipinnate (mainly from Africa and
Asia).
Phyllodenous Acacia species
The majority of Australian Acacia spp. lack true leaves.
Instead, they exhibit a characteristic known as phyllodony where the petiole is
expanded and flattened. The phyllode has the ability to photosynthesize and for
all practical purposes, it is the equivalent of a leaf. The Australian
exceptions are from the Section Botrycephalae, which maintain bipinnate foliage
until maturity and do not develop phyllodes.
Vercoe (1987) noted that species of Acacia and other genera from
the dry tropics of Australia had potential in the semi-arid zones of Africa for
fuelwood and erosion control. If their usefulness as fodder could also be
established, this would enhance their overall value. Certainly the value of
trees such as A. aneura as drought reserve feed is well established and
recognized by pastoralists in Australia. Unfortunately, success in establishing
A. aneura in Africa has been limited. However, a number of other species appear
to show potential, notably A. holosericea, A. trachycarpa and A. coriacea. Other
species that have been studied include A. salicina and A. cyanophylla, although
the phyllodes of the latter contain high tannin levels (Vercoe, 1987; Goodchild
and McMeniman, 1987; Reed et al., 1990), which can reduce both the palatability
and the feeding value. Unlike African and Asian species, Australian Acacia spp.
are evergreen and
unarmed.
Bipinnate Acacia species
There are about 200 bipinnate Acacia species in the New World
and a further 150 species in Africa and Asia. All 129 African species possess
bipinnate leaves. There are other physical and physiological characteristics
which separate them from their Australian relatives, the Botrycephalae. This
group, the best known of which is A. mearnsii (black wattle), grown throughout
the subtropical world for the extraction of tannin, is confined to the cooler
and moister parts of south eastern Australia. They are not noted as valuable
fodder trees (Boland, 1987). Most bipinnate Acacia spp. are armed with thorns of
various shapes and sizes, some of which are modified stipules at the base of the
leaves. Many exhibit transient periods of defoliation in the dry season and have
been described as deciduous. Before the start of the rains, there is a often a
flush of new growth of leaves. Although the stimulus for this activity is
uncertain, it may be due to
temperature.
Fodder characteristics
Fodder yield
Information on leaf yield of African bipinnate Acacia spp. is
limited, and comparative data with introduced Australian species, even more so.
The work of Cossalter (1986) compared species from the two regions, when grown
in Senegal at uniform spacings of 3 x 3 m (Table 1). The results, as means of
20, or 25 individual trees aged between 18-23 months, demonstrated the fodder
potential of the Australian Acacia species when compared with African ones. A.
holosericea is perhaps the most frequently planted Australian Acacia in
development projects because of its superior yield and because it retains a
large phyllode biomass during the dry season, while African species shed their
leaves during this period (Cossalter, 1986). There is some uncertainty about the
palatability of A. holosericea: early trials indicate that only dry phyllodes
are accepted. The fresh phyllodes of A. trachycarpa and A. tumida, however, were
readily eaten by both cattle and sheep (Hamel, 1980).
Table 1 - Total biomass production (phyllodes, leaves and
wood) of Acacia spp.
|
Origin phyllodes |
Species(kg/tree) |
Dry leaves, (g/tree) |
Wood |
|
Australia |
A. Holosericea |
2660 |
12.03 |
|
Australia |
A. trachycarpa |
1993 |
10.65 |
|
Australia |
A. tumida |
2600 |
8.69 |
|
Australia |
A. trachycarpa |
1700 |
3.73 |
|
Africa |
A. senegal |
60 |
9.92 |
|
Africa |
A. seyal |
208 |
9.39 |
|
Africa |
A. tortilis |
130 |
5.89 |
Source: Cossalter (1986)
When assessing the fodder value of African Acacia spp., the
importance of the fruit and pods cannot be ignored. This fraction may be of more
value than the foliage in some cases. A major problem in estimating the yield of
fruit is the variation in individual trees between years. Trees may bear a heavy
crop one year and little the next. The fruits of F. albida are relished by game,
cattle and small ruminants and NAS (1979) suggests that yields of 125-135 kg are
possible from a single tree. When ripe, the fruit can be harvested and stored
for future use, although there is a tendency for the pods to shatter and
disintegrate.
The bark of A. seyal is particularly valuable as a fodder,
although complete removal will result in the death of the tree. Skerman et al.,
(1988) suggested that the bark, which is extensively used for feeding cattle and
small ruminants in the dry season, was the most valuable part of the plant.
Cattle could consume up to 5.5 kg bark/day, sufficient for maintenance and the
production of 4.5 litres of milk.

Table 2 - Mean proximate and fibre
analyses of Acacia leaves, phyllodes, fruits and
seeds
Nutritive value
In Table 2, the mean proximate and fibre analyses of leaves,
phyllodes and fruits of Acacia spp. are summarized from data of individual
species (Appendix 1). Absolute values should be treated with caution since there
is considerable variation in the literature. For example, the mean crude protein
(CP) level of Acacia leaves is 18.9% of dry matter (DM), with a reported range
of 6.5-42.8%. However, the data provide an indication of the nutritive value and
in general both leaves and fruits of all species are valuable sources of
protein, particularly when compared with mature grasses, where CP levels in
forage dry matter (DM) during the dry season can fall to 3-4% or less, well
below the 7% CP which is suggested as a lower limit for fibre digestion in
cattle (Van Soest, 1982).
While the crude fibre (CF) levels of Acacia are relatively high
(15-40%) compared with other fodder tree species such as Leucaena leucocephala
and Gliricidia septum, they are still markedly lower than for tropical grasses
and common crop residues. Even the fruit, consisting of the dry dehiscent pod
and the seeds contained in it, are a valuable feed source, with moderate CF
(25%) and high CP levels (15%). The phyllodenous Acacia spp. tend to have
marginally higher CF and lower CP levels than bipinnate species (Appendix 2)
though the available data consist of fewer observations. The lower nutritive
value is offset by both the higher biomass yield and the retention of the
phyllodes throughout the dry season, when bipinnate species tend to shed their
leaves.
Information on the digestibility of DM and organic matter (OM)
of Acacia leaves and phyllodes is limited but the available data indicate that,
with sheep, they are relatively low and in the range of 40-605 to respectively
(Table 3). The digestibility of phyllodes appears to be lower than that of
bipinnate leaves. Goodchild and McMeniman, (1987) suggested that the low fibre
digestibility could be associated with the high lignin content of the cell wall,
since fibre digestibility is inversely related to the lignin content of the
fibre fraction.
Information on the digestibility of DM and organic matter (OM)
of Acacia leaves and phyllodes is limited but the available data indicate that,
with sheep, they are relatively low and in the range of 40-60% respectively
(Table 3). The digestibility of phyllodes appears to be lower than that of
bipinnate leaves. Goodchild and McMeniman, (1987) suggested that the low fibre
digestibility could be associated with the high lignin content of the cell wall,
since fibre digestibility is inversely related to the lignin content of the
fibre fraction.
While the digestibility of the whole fruit (seed and pod) is
higher than that of the leaves and phyllodes, it can be adversely affected by
the proportion of seeds that pass undigested through the gut of the animal.
Tanner et al., (1990) reported that 46% of A. tortilis seeds were voided in the
faeces of sheep. Similarly, Goodchild and McMeniman (1987) noted that when
feeding yearling cattle with seeds of A. tortilis subsp. spirocarpa, 12% passed
unchanged through the gut. Legume seeds are physically hard and notoriously
resistant to digestive juices, although the small size of Acacia seed may be a
contributory factor in their passage through the gut (Tanner et al., 1990).
The digestibility of the leaf and fruit fraction can also be
depressed by the presence of polyphenolic compounds, including
tannins.
Anti-nutritive factors
Acacia spp. have acquired a variety of physical and chemical
defence mechanisms aimed at reducing the palatability and nutritive value of the
plant to predators including both insects and animals.
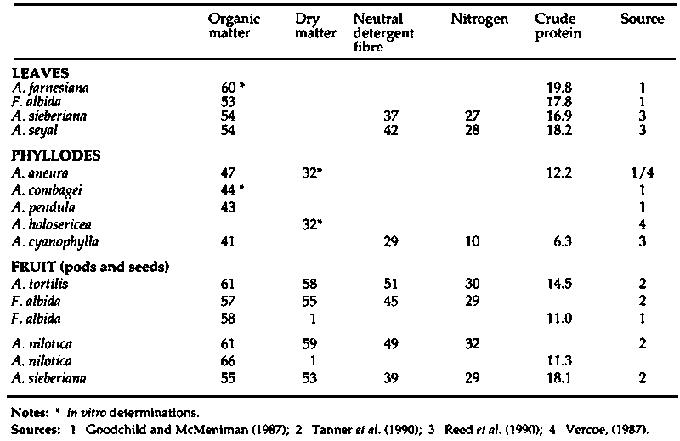
Table 3 - In vivo
digestibility (%) of Acacia forage
Perhaps the most obvious physical characteristic is the spiny
armature of the African species, which evolved as a defence against large
herbivores. A. mellifera subsp. detinens derived its common name (wait-a-bit
tree) from the pairs of small hooks on the branches. A. tortilis is protected by
two types of thorns, long white stipular spines and small curved hooks, a
formidable protection against animals. Coe and Beentje (1991) suggested that as
thorns restrict the availability of forage, a critical level is reached below
which the herbivore no longer finds it nutritionally efficient to seek shoots
between the spines, since this invariably involves movement between each bite.
There is also evidence that in A. drepanolobium, thorns on branches which are
regularly browsed by animals are significantly longer than those on undefoliated
branches (Young, 1987).
Tree size is a form of physical protection that can be harnessed
as a desirable characteristic in certain agroforestry systems. Domestic animals
are unable to browse tall trees which then maintain a standing reserve of
foliage which can be made available by strategic lopping at critical periods
when other feed is scarce.
A fascinating defence mechanism of the genus is the symbiotic
relationship with ants known as myrmecophily, where the plant lives in
association with an ant colony. In the tropical American species A. cornigera,
association with ants from the genus Pseudomyrmex is a notable example of
myrmecophily. The trees have modified, swollen spines or galls, which provide
shelter for the ants, and extrafloral nectaries and modified leaflet tips which
provide food. The ants, in return, protect the trees from insects and other
predators by swarming to attack them when the tree is disturbed (Janzen, 1966).
In Africa, A. drepanolobium is commonly known as the 'whistling thorn', a name
derived from the noise of the wind blowing into the holes of the galls that have
been occupied by ants.
Many Acacia spp. produce a range of potentially toxic compounds
which prevent herbivores from eating the foliage, although it is thought that
the chemicals originally evolved as defence mechanisms against insect and fungal
attack. Some members contain specific anti-nutritive factors, for example, A.
georginae has been implicated in heavy stock losses in Australia due to its
content of fluoracetic acid (D'Mello, 1992). In general terms the genus contains
two major groups of anti-nutritive factors, cyanogenic glycosides and
polyphenolic
compounds.
Cyanogenic glycosides
When cyanogenic substrates are broken down under the action of a
hydrolyzing enzyme, hydrogen cyanide (HCN) is formed. Plants that contain both
cyanogenic glycosides and an endogenous, hydrolytic enzyme are the most
dangerous but broad-spectrum enzymes may be present in other fractions of the
diet. Symptoms of HCN poisoning include respiratory dyspnoea, intense red
conjunctive, frothing at the mouth, bloat, a staggering gait, convulsions and
violent death. On post-mortem examination, a characteristic almond smell is
often noted in the contents of the gut.
In Asia, there are a number of reports of HCN poisoning of
livestock when fed on the pods and leaves of A. Ieucophloea (Gupta and Naurival,
1966; Prasad et al., 1977; Vihan and Panwar, 1987). In South Africa, Steyn and
Rimington (1935) reported the presence of cyanogenic glycosides in A.
haematoxilon, A. Iasiopetala, A. tortilis subsp. heteracantha, A. robusta and A.
hebeclada subsp. hebeclada. They found higher concentrations of HCN from leaves
and immature pods than from mature pods. In A. robusta, the glycoside was
present without the corresponding hydrolytic enzyme. They isolated the
cyanogenic compound acacipetalin from A. Iasiopetala and A. hebeclada. Siegler
et al., (1975) isolated a further compound, dihydroacipetalin, from the leaves
and shoots of A. sieberiana var. woodii.
Toxicity appears only in sporadic outbreaks, being dependent on
a number of factors, including the species and variety of Acacia, the rate of
ingestion by the animal and the availability of alternative feeds. It is more
likely to occur during periods of drought or feed scarcity when hungry animals
consume large quantities of feed in a short period of time. Access to cold water
during feeding appears to promote the release of HCN from the substrates.
Practical measures can be taken to reduce the risk of poisoning such as
restricting the intake, particularly of leaves and immature pods, separating the
time or place of feeding and watering, and feeding dangerous material with
molasses or sulphur. These substances combine with HCN to form harmless
cyanhydrin and sulphocyanides respectively (Steyn and Rimington,
1935).
Phenolic compounds
There are numerous reports of the presence of polyphenolic
compounds in the various components of plants of the genus Acacia. From the
standpoint of nutrition, the most important of these are the tannins, compounds
with molecular weights in excess of 500, which are able to form complexes with,
and to precipitate proteins (McLeod, 1974; Mangan, 1988). Hydrolyzable tannins
are readily broken down by acids, bases and enzymes to form sugars and phenolic
carboxylic acid and are therefore easily degraded in the gut. In contrast,
condensed tannins are not degraded and so are available to form complexes with
proteins, resulting in reduced feed intake and digestibility. McNaughton (1987)
suggested that at levels above about 5%, increasing the tannin content in a diet
reduced its palatability.
Reed et al., (1990) assessed the effects of polyphenolics on
intake, growth, digestibility and nitrogen utilization by sheep. Three Acacia
forages, (phyllodes of A. cyanophylla, leaves of A. seyal, and fruit of A.
sieberiana), all containing high levels of polyphenolics, were compared with
three protein supplements and another fodder tree Sesbania sesban which is low
in polyphenolics. All diets were fed in combination with teff straw (Eragrostis
abyssinica). The diets containing Acacia led to the highest levels of feed
refusal by the sheep their and lowest intake of teff straw. Fibre digestibility
was lower for these diets and the tannins complexed with proteins, resulting in
higher levels of faecal nitrogen.
A complementary study by Tanner et al., (1990) reported
significant differences in the growth rates of sheep supplemented with fruits
from four different Acacia spp. (A. tortilis, F. albida, A. nilotica and A.
sieberiana) even though there were no significant differences in total nitrogen
intake. The tannin-rich fruits, particularly of A. sieberiana and A. nilotica,
reduced feed intake and the efficiency of nitrogen utilization by forming
complexes with proteins which again resulted in a shift of nitrogen excretion
from urine to faeces. The growth rates of sheep fed the diets low in
polyphenolics (F. albida and A. tortilis), were not significantly different from
sheep supplemented with noug meal (Guizotia abyssinica), indicating their
suitability as dietary supplements.
The literature abounds with conflicting reports on the
suitability of Acacia spp. for livestock feed.
Dumancic and Le Houerou (1981) considered the phyllodes of A.
cyanoophylla to be sufficient to meet the protein maintenance requirements of
sheep, while Reed et al., (1990) reported a negative nitrogen balance for
animals fed on this legume. Predictions of the productive responses of livestock
are problematical and uncertain. There are a number of possible explanations for
the conflicting reports:
· the chemistry of tannins is
complex and analytical results do not always correlate with biological effects,
e.g. nitrogen digestibility;
· a single feed may contain more
than one antinutritive factor. A. nilotica is known to have both tannins and
catechin gallates (Mueller-Harvey et al., 1987);
· the concentration and type of
phenolic compound both vary with season and in response to stimuli such as
defoliation;
· there is much variation
between both species and provenance in tannin content, the effect being both
genetic and
environmental.
Management
Where the principal use of Acacia spp. is for browsing by
livestock, little is done to manage the resource, except through some measure of
defoliation control by herding the animals. High branches are lopped during
times of feed shortage, particularly in the dry season. At other times of the
year, the trees are subjected to uncontrolled browsing, often by both wild and
domestic animals.
Many legumes produce seed which has a hard, impermeable seed
coat. Such seed will not germinate until moisture is able to penetrate it. This
characteristic, known as 'hard seededness', prevents the simultaneous
germination of the entire stock of seed in the soil at the start of the wet
season. This adaptation favours the long-term survival of the population where
rainfall is unpredictable. It is, however, a disadvantage when attempting to
establish new stands from seed, particularly in areas with a short growing
season. Acacia shows extreme variability in the degree of hard seededness, both
between and within species; the proportion of hard seed in a sample depends on
the environmental conditions during plant growth, the maturity of the seed at
harvest and the length of time it has spent in storage. In general, Australian
species respond to seed scarification. Recommended treatments (Doran and Gunn,
1987) include:
· manual nicking of the seed
coat. Because of the amount of labour involved, this is suitable only for seed
for germination tests, or for valuable research lots;
· placing the seed in boiling
water for one minute. This is recommended for many hard-coated seed lots;
· placing the seed in hot water
(80-90°C) for one minute. This is recommended where sensitivity to boiling
water is suspected.
In Central Australia where annual rainfall is about 300 mm,
native Acacia spp. naturally regenerate after exceptionally high rainfall at
populations of some 5000 stems/ha, and many species produce multiple stems.
Subsequent fires and drought will thin the stand to 100-400 stems/ha. Under
these conditions,
the optimum stocking level for planted stands may be in the
region of 1000 seedlings/ha, followed by thinning as the stand matures.
Alternatively, where thinning is not practical, an initial stand of about 400
trees/ha would be appropriate (Kube, 1987). The seed is scarified in hot water
(80°C) and germinated in trays. Seedlings are transplanted after a few days
into individual tubes 5 cm in diameter by 15 cm deep, filled with a potting
mixture of equal parts of coarse sand, fine sand or sandy loam and peat moss.
Treatment with fungicide is usually necessary and the seedlings are grown in
decreasing shade for about three months before planting out. Supplementary
irrigation is applied for 6-12 months, depending upon rainfall. Growth is often
rapid, many species attaining heights of 2 m in about two years.
In Tanzania, A. mearnsii plantations have been established for
fuelwood, charcoal and building poles by direct seeding scarified seed at rates
of 1.22.4 kg/ha in rows 1.8 m apart in fully cultivated soil (Kessy, 1987). The
major problem with this species has been its susceptibility to competition from
weeds. Harvesting has been on an 8-12 year cycle and natural regeneration has
taken place after the original stand has been clear felled and
burned.
Alternative uses
While Acacia is the largest of the mimosoid genera and the
second largest of all the legume family, only about 75 members have economic
value, and of these only about 50 are actually cultivated. Nevertheless, they
are used in various ways, as described by Allen and Allen, (1981) and Boland,
(1987).
Many Acacia spp. form spiny, slow-growing scrub which provides
protection against soil erosion caused by wind and rain. Their tolerance of
harsh conditions makes them valuable for stabilization and revegetation of
difficult sites. A. armata, A. glaucescens, A. pycnantha and A. suaveolens
tolerate salt spray and so can be used to stabilize coastal sand dunes. A.
decurrens is planted along roadsides in Australia and on denuded hills in
Rwanda. A. mearnsii and A. dealbata are used to control gulley and hillside
erosion in New Zealand.
Acacia is an important source of fuelwood or charcoal in arid
regions. A. mearnsii is used for both domestic fuel and tobacco curing in
Indonesia (NAS, 1980). It also provides poles for mining and agriculture in many
parts of the world.
As timber, Acacia wood is extremely durable and although it is
hard to work, it finishes well. Smaller species provide wood which is used to
make implement handles, pipes and furniture, while larger species are used for
paneling, boats and musical instruments. Hawaian mahogany (A. koa) was once
prized for making surfboards, canoes and bowls while A. segal and A. tortilis
were the principal trees of the Arabian Desert region. A. glaucescens and A.
melanoxylon are major timber trees today. A. dealbata and A. mearnsii are used
for pulping in Australia and Japan, while the latter species is used in the
production of rayon in South Africa and India.
A. mearnsii is the world's most important source of vegetable
tannin, 35-40% of the air-dry weight of bark is extracted for leather
manufacture. Yields in excess of 15% are also obtainable from other species such
as A. albida and A. cyanophylla. Wood adhesives and polyurethane foams can be
prepared from bark extracts of A. mearnsii, and used as substitutes for
petroleum products.
Gum arable, an important emulsifier, protective colloid,
adhesive and binder, is produced in commercially exploitable quantities by A.
arabica, the principal historical source, and more recently by A. senegal and
some 20 other species of Acacia. It is used in foods, pharmaceuticals,
cosmetics, adhesives, paints, polishes and inks and in lithography, photography,
textile-sizing and other processes.
Species such as A. dealbata, known as mimosa by florists, A.
albida and A. tortilis, are valued as ornamentals while A. farnesiana and A.
dealbata are used in the production of base oils for perfumes. In Australia,
several species are valuable sources of pollen in
apiculture.
References and further reading
AHN, J.H., ROBERTSON, B.M., ELLIOT, R., GUTTERIDGE, R.C. and
FORD, C.W. (1989) Quality assessment of tropical browse legumes: tannin content
and protein degradation. Animal Feed Science and Technology, 27: 147-156.
ALLEN, O.N. and ALLEN, E.K. (1981) The Leguminosoe: a Source
Book of Characteristics, Uses and Nodulation. Madison, USA: University of
Wisconsin Press.
BATE-SMITH, E.C. (1973) Haemanalyis of Tannins: The Concept of
Relative Astringency. Phytochemistry, 12: 907-912.
BENTHAM, G. (1864) Flora Australiensis vol.2. London: Reeve and
Co.
BOLAND, D. J. (1987) Genetic resources and utilisation of
Australian bipinnate acacias (Botrycephalae). In: Australian Acacias in
Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop
held at Gympie, Australia, August, 1986. ACIAR Proceedings No.16. pp. 29-37.
BUTLER, L.G. (1982) Relative degree of polymerization of sorghum
tannin during seed development and maturation. Journal of Agriculture and Food
Chemistry, 30: 1090-1094.
COE, M. and BEENTJE, H. (1991) A Field Guide to the Acacias of
Kenya. Oxford, UK: Oxford University Press.
COSSALTER, C (1986) Introduction of Australian Acacias into Dry
Tropical West Africa. Forest Ecology and Management, 16: 367-389.
D'MELLO, J.P.F. (1992) Chemical Constraints to the use of
tropical legumes in animal nutrition. An�mal Feed Science and Technology, 38:
237-261
DORAN, J.C. and GUNN, B.V. (1987) Treatments to promote seed
germination in Australian Acacias. In: Australian Acacias in Developing
countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at
Gympie, Australia, August 1986. ACIAR Proceedings No. 16. pp.
57-63.
DUMANCIC, D. and LE HOUEROU, H.N. (1981) Acacia cyanophylla
Lindl. as a supplementary feed for small stock in Libya. Journal of Arid
Environments, 4: 161 -167.
GOHL, B. (1981) Tropical Feeds: Feed Information Summaries and
Nutritive Values. FAO Animal Production and Health Series No. 12. Rome: FAO.
GOODCHILD, A.V. and McMENIMAN, N.P. (1987) Nutritive value of
Acacia foliage and pods for animal feeding. In: Australian Acacias in Developing
Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop held at
Gympie, Australia, August 1986. ACIAR Proceedings No.16. pp. 101-106.
GUPTA, I. and NAURIYAL, M.M. (1966) Acacia leucophloea Willd
(raunja) poisoning in livestock. Indian Veterinary Journal, 43: 538-543.
HAGERMAN, A.E. and BUTLER, L.G. (1981) The specificity of
proanthocyanidin-protein interactions. Journal of Biological Chemistry, 256:
4494-4497.
HAMEL, O. (1980) Acclimatization and utilization of
phyllodineous acacias from Australia in Senegal. In: Browse in Africa; The
current state of knowledge. LE HOUEROU, H.N. (ed.) International Livestock
Centre for Africa: Addis Ababa. pp. 361-373.
JANZEN, D.H. (1966) Coevolution of mutualism between ants and
acacias. Evolution, 20: 249-275.
KESSY, B.S. (1987) Growth of Australian Acacias in Tanzania. In:
Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.) Proceedings,
International Workshop held at Gympie, Australia, August 1986. ACIAR Proceedings
No. 16. pp. 123-125.
KUBE, P.D. (1987) Growth rates, establishment techniques and
propagation of some Central Australian Acacias. In: Australian Acacias in
Developing Countries. TURNBULL, J.W. (ed.) Proceedings, International Workshop
held at Gympie, Australia, August 1986. ACIAR Proceedings No. 16. pp. 77-80.
KUMAR, R. and VAITHIYANATHAN, S. (1990) Occurrence, nutritional
significance and effect on animal productivity of tannins in tree leaves. Animal
Feed Science and Technology, 30: 21-38.
LE HOUEROU, H.N. (1980) The role of browse in the Sahelian and
Sudanian zones. In: Browse in Africa; The current state of knowledge. LE
HOUEROU, H.N. (ed.) International Livestock Centre for Africa: Addis Ababa. pp.
83-100.
LOCK, J.M.(1989) LegumesofAfrica: ACheckList. Kew: Royal Botanic
Gardens.
MAKKAR, H.P.S., DAWRA, R.K. and SINGH, B. (1988) Changes in
tannin content, polymerisation and protein precipitation capacity in oak
(Quercus incana) leaves with maturity. Journal of Science of Food and
Agriculture, 44: 301-307.
MANGAN, J.L. (1988) Nutritional effects of tannins in animal
feeds. Nutrition Research Reviews, 1: 209-231.
MASLIN, B.R., CONN, E.E. and DUNN, J.E. (1987) Cyanogenic
Australian species of Acacia: A preliminary account of their toxicity potential.
In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.)
Proceedings, International Workshop held at Gympie, Australia, August 1986.
ACIAR Proceedings No.16. pp. 107-111.
McLEOD, M.N. (1974) Plant tannins - their role in forage
quality. Nutrition Abstracts and Reviews, 44: 803-815.
McNAUGHTON, S.J. (1987) Adaption of herbivores to season changes
in nutrient supply. In: Nutrition of Herbivores. HACKER, J.B. and TERNMOUTH,
J.H. (eds). Sydney, Australia: Academic Press. pp. 391-408.
MUELLER-HARVEY, I., REED, J.D. and HARTLEY, R.D. (1987)
Characterisation of phenolic compounds, including flavonoids and tannins, of ten
Ethiopian browse species by high performance liquid chromatography. Journal of
the Science of Food and Agriculture, 39: 1-14.
NAS (1979) Tropical Legumes: Resources for the Future.
Washington DC: National Academy of Sciences.
NAS (1980) Firewood Crops. Washington DC: National Academy of
Sciences.
PEDLEY, L. (1986) Deprivation and dispersal of Acacia
(Leguminosue) with particular reference to Australia, and recognizing Senegalia
and Racospenna. Botanical Journal of the Linnean Society, 92: 219-54.
PEDLEY, L. (1987) Australian Acacias: Taxonomy and
Phytogeography. In: Australian Acacias in Developing Countries. TURNBULL,
J.W.(ed.) Proceedings, International Workshop held at Gympie, Australia, August
1986. ACIAR Proceedings No 16. pp. 11-16.
PRASAD, J., SINGH, A.P. and REKIB, A. (1977) Hydrocyanic acid
poisoning in grazing sheep and goat on Acacia leucophloca (reunja). Indian
Veterinary Journal, 54: 748-751.
REED, J.D., HORVARTH, P.J., ALLEN M.S. and VAN SOEST, P.J.
(1985) Gravimetric determination of soluble phenolics including tannins from
leaves byprecipitation with trivalent ytterbium. Journal of the Science of Food
and Agriculture, 36: 255-261.
REED, J.D., SOILER, H., and WOODWARD, A. (1990). Fodder tree and
straw diets for sheep: intake, growth, digestibility and the effects of
phenolics on nitrogen utilisation. Animal Feed Science and Technology, 30: 3950.
SEDDON, H.R. and KING, R.O.C. (1930) As to the fatal dose for
sheep of cyanogenetic plants containing Sambunigrin and Prunasin. Journal
Council Scientific and Industrial Research Australia, 3: 14-24.
SIEGLER, D.S, BUTTERFIELD, C.S., DUNN, J.E. and CONN, E.E.
(1975). Dihydroacacipetalin - a new cyanogenic glycoside from Acacia sieberiana
var woodii. Phytochemistry, 14: 1419-1420.
SKERMAN, P.J., CAMERON, D.G. and RIVEROS, F. (1988) Tropical
Forage Legumes. 2nd. edn. Rome: FAO.
STEYN, D.G. and RIMINGTON, C. (1935). The occurrence of
cyanogenetic glucosides in South
African species of Acacia. Onderstepoort Journal of Veterinary
Science and Animal Industry, 4(1): 51-73.
TANNER, J.C., REED, J.D. and OWEN, E. (1990). The nutritive
value of fruits (pods with seeds) from four Acacia spp. compared with extracted
noug (Guizotia abyssinica) meal as supplements to maize stover for Ethiopian
highland sheep. Animal Production, 51: 127-133.
TAUBERT, P. (1894) Die Naturlichen Planzenfamilie. Leipzig:
Wilhelm Engelmann.
TERBLANCE, M., PIENAAR, J.G., BIGALKE, R. and VAHRMEYER, J.
(1967) Acacia nilotica (L.) Del. subsp. kraussiana (Berth) Brenan. as a
poisonous plant in South Africa. Journal South African Veterinary Medical
Association, 38(1): 57-63.
VAN HOVEN, W. (1985) The tree's secret weapon. South African
Panorama, 30(3): 34-37.
VAN SOEST, P.J. (1982) Nutritional Ecology of the Ruminant.
Corvallis, Oregon, USA: O & B Books.
VERCOE, T.K. (1987) Fodder potential of selected Australian tree
species. In: Australian Acacias in Developing Countries. TURNBULL, J.W. (ed.)
Proceedings, International Workshop held at Gympie, Australia, August 1986.
ACIAR Proceedings No.16. pp. 95-100.
VIHAN, V.S. and PANWAR, H.S. (1987) A note on toxicity of Acacia
leucophloea (Rajma) in sheep. Indian Journal Animal Research, 21(1): 53-55.
WILSON, A.D. (1969). A review of browse in the nutrition of
grazing animals. Journal of Range Management, 22: 23-28.
WOODWARD, A. and REED, J.D. (1989) The influence of
polyphenolics on the nutritive value of browse: A summary of research conducted
at ILCA. ILCA Bulletin 35, Addis Ababa: International Livestock Centre for
Africa.
YOUNG, T.P. (1987) Increased thorn length in Acacia
drepanolobium - an induced response to browsing. Oecologia 71(3):
436-438.
Appendix 1
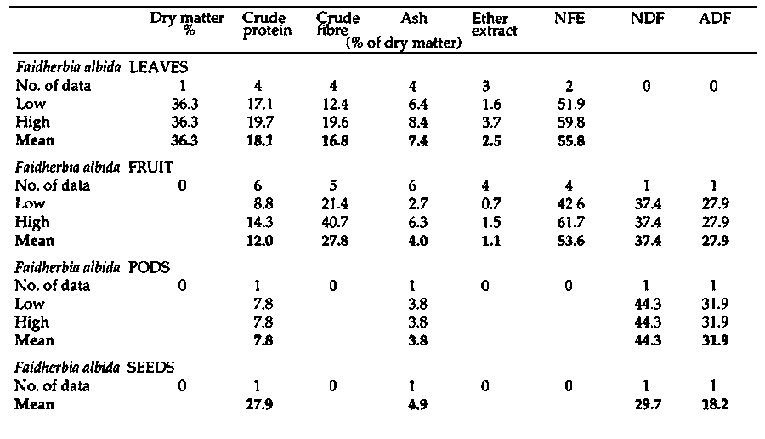
Primate and fibre analyses of
bipinnate Acacia spp.
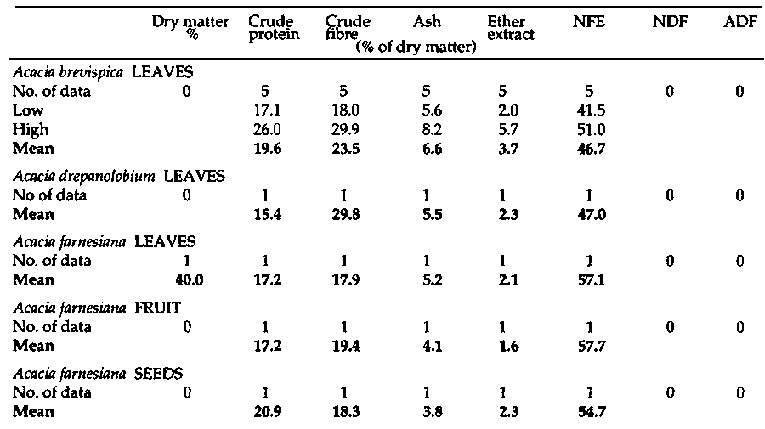
Primate and fibre
analyses of bipinnate Acacia spp. (continued 1)
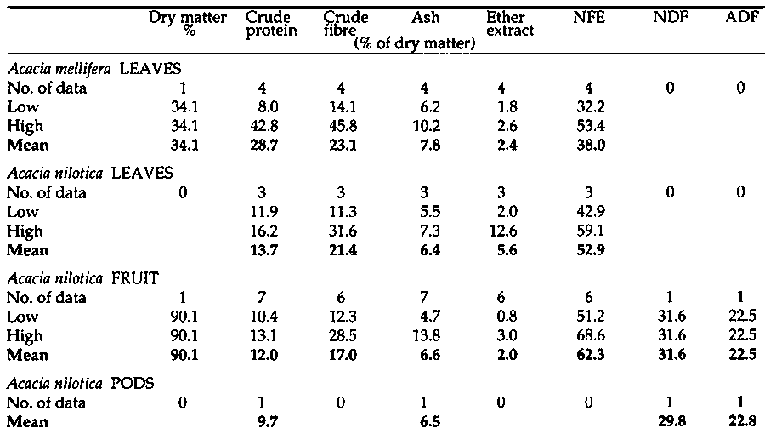
Primate and fibre analyses of bipinnate Acacia spp. (continued
2)
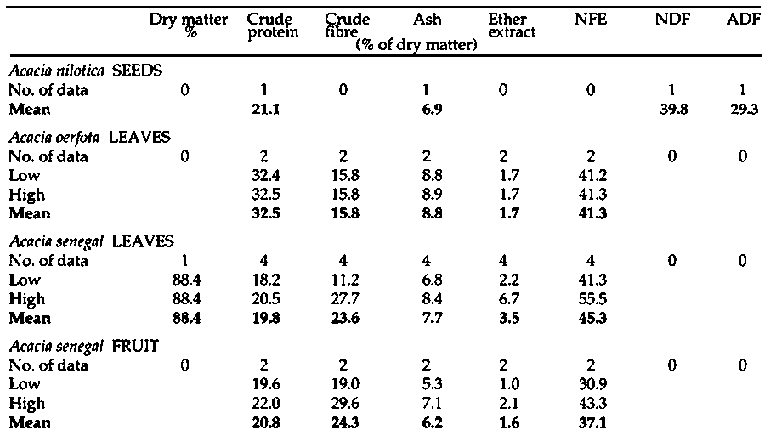
Primate and fibre analyses of bipinnate
Acacia spp. (continued 3)
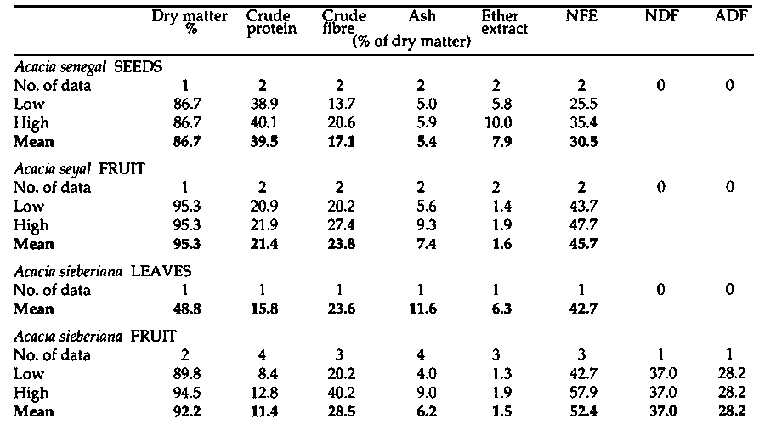
Primate and fibre
analyses of bipinnate Acacia spp. (continued 4)
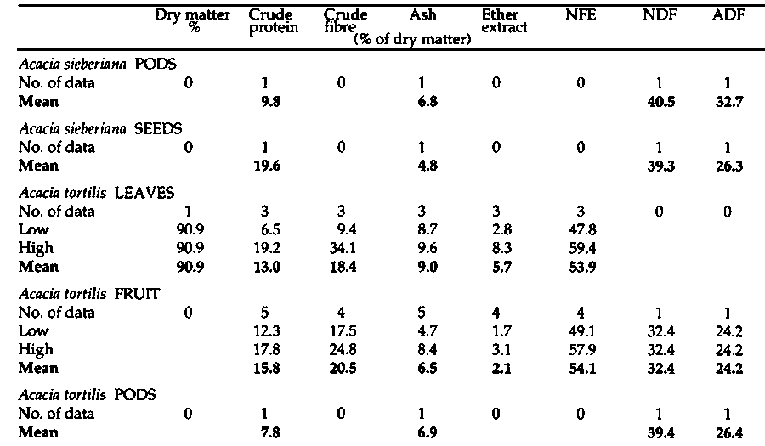
Primate and fibre analyses of bipinnate Acacia spp. (continued
5)
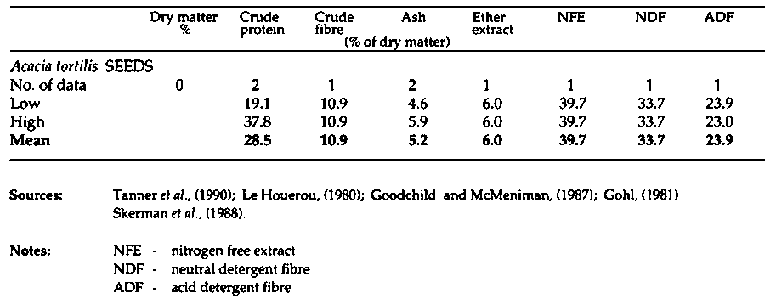
Primate and fibre analyses of bipinnate
Acacia spp. (continued
6)
Appendix 2
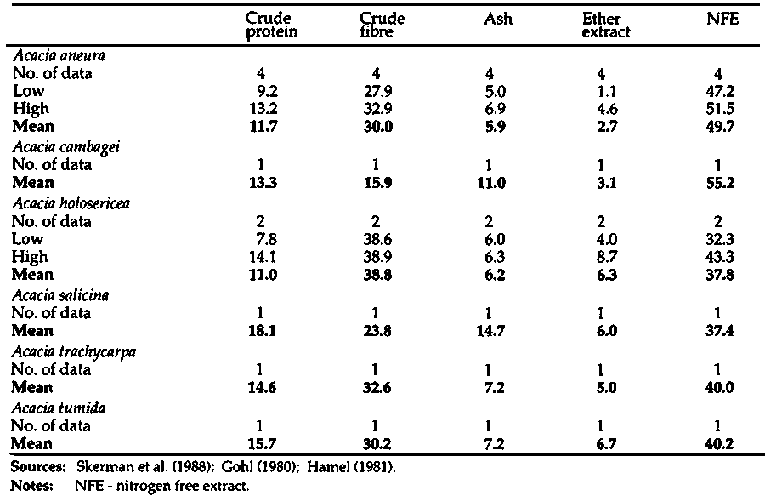
Proximate analyses of Acacia
phyllodes
